
Fúze chromozomů
Chromozomy jsou klasifikovány na základě umístění centromery (zúžené místo na chromozomu, kde jsou sesterské chromatidy po replikaci navzájem spojeny). Když je centromera uprostřed chromozómu a obě ramena mají stejnou délku, je chromozóm označován jako metacentrický chromozóm. Pokud se centromera nalézá poblíž konce chromozomu, pak se chromozóm nazývá akrocentrický. Veškeré změny ve struktuře chromozómu je možné odhalit pomocí zvláštního proužkovacího barvení.
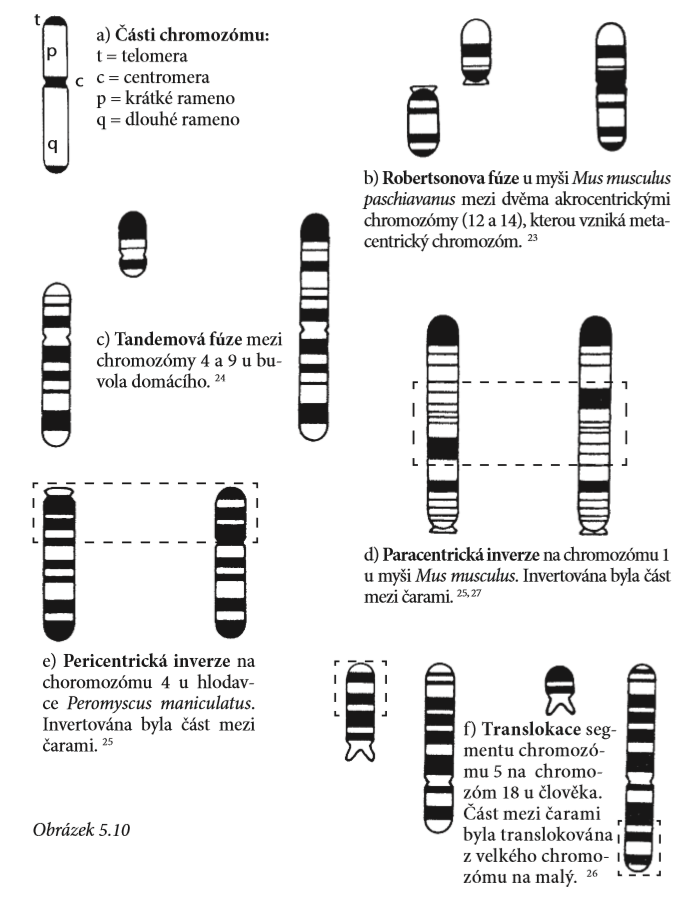
Změny na chromozomech mohou zahrnovat změny v počtu chromozomů, počtu ramen i jiné změny vyvolané translokací (přesunem segmentu chromozomu do jiného místa), vynechání, duplikace, inverze i drastické změny uspořádání. Chromozómy mohou občas fúzovat (spojovat se), přičemž se vytvářejí mnohem delší chromozómy, nebo se mohou v centromeře rozdělit, čímž se vytvoří dva kratší chromozómy. Jedna takováto změna uspořádání je označována jako Robertsonovo přeuspořádání a je výsledkem buď splynutí dvou centromer do jedné, nebo rozdělení jedné centromery na dvě. Tandemová fúze je naopak splynutí dvou chromozómů, při němž se jeden konec chromozómu spojí s koncem nebo centromerou jiného chromozómu (viz obrázek 5.10). Při těchto rekombinacích je informace přeskupena, ale srovnání mezi proužkováním chromozómů u příbuzných druhů s různou chromozómovou strukturou ukazuje, že informace je stále stejná. Je jen jinak uspořádaná. Navíc platí, že typ změny v uspořádání, k němuž dochází u různých živočichů, je specifický pro určitou skupinu, a jeden typ přeskupení se tedy nemusí nutně vyskytovat u jiné.
Podívejme se na způsoby, kterými může docházet ke změně ve struktuře a počtu chromozomů:
Robertsonovo přeuspořádání
Jedna takováto změna uspořádání je označována jako Robertsonovo přeuspořádání a je výsledkem buď splynutí dvou centromer do jedné, nebo rozdělení jedné centromery na dvě.
Robertsonova fúze mění počet chromozomů, ale ne počet ramen. Když se chromozomy uspořádávají během meiózy 1, metacentrický chromozóm se stěhuje ke dvěma akrocentrickým chromozómům. Příkladem může být myš domácí (Mus musculus), která má 40 chromozómů. V Alpách žije populace myší, u kterých bylo zjištěno jen 22 chromozómů. Tato populace se také mírně morfologicky liší od normální myši domácí a je zařazena do samostatného druhu Mus paschiavanus.
Byly objeveny i jiné populace s počtem chromozómů mezi 22 a 40, přesto však počet ramen zůstává stejný a proužkování ukazuje, že geny jsou homologické. Z hlediska jejich vztahu je zjevné, že všechny tyto různé druhy patří do stejné skupiny.
Tandemová fúze
Tandemová fúze je naopak splynutí dvou chromozómů, při němž se jeden konec chromozómu spojí s koncem nebo centromerou jiného chromozómu. Při tandemové fúzi se mění počet ramen a počet chromozómů.
Tandemová fúze byla zjištěna u několika druhů antilop, u kterých došlo k fúzi chromozómu kódujícího pohlaví s jedním z autozómů. K tomuto jevu dochází jen zřídka, a můžeme proto předpokládat, že organismy zřejmě měly společného předchůdce.
Antilopy, u nichž se projevuje tato fúze, se liší velikostí od antilopy losí (největší z antilop) až po menší druhy, jako sitatungu a lesoně jižního. Všichni mají ovšem i stejné rysy, jako například podobný tvar rohů a pruhů na těle, které mohou být velice výrazné (například u antilopy bongo), nebo méně nápadné (jako u antilopy losí). U druhů, u kterých se objevuje tato fúze: antilopa losí, bongo, kudu velký, kudu malý, lesoň jižní, sitatunga a nilgan pestrý (antilopa žijící v Indii), došlo k fúzi (sloučení, spojení) chromozómu y a autozómu.
Tandemovou fúzi najdeme i u malajského vodního buvola a asijského říčního buvola.
Další zajímavý příklad fúze nalézáme u asijských jelenů. U muntžaka červeného (Muntiacus muntjac) má samice jen 6 chromozómů, zatímco samec 7 (nejmenší počet chromozómů u savce). U jiného druhu této podčeledi, muntžaka malého (Muntiacus reevesi), mají samci i samice 46 chromozómů.
Výzkum proužkování ukazuje, že u obou druhů je přítomen stejný genetický materiál. Chromozómy muntžaka červeného však prošly fúzí, přičemž se vytvořily velice dlouhé chromozómy. Ani v tomto případě nebyla přidána nová informace, jen se jinak uspořádala, a tím umožnila jiné vyjádření genů a vzrůst počtu variací – stejně jako je možné na tomtéž klavíru zahrát spoustu různých melodií, a přesto hudba stále zůstává klavírní hudbou.
Translokace
Translokace genu může vést ke snížení plodnosti. U lidí může dojít ke vzniku tzv. Downova syndromu, pokud je část 21. chromozómu translokována na jiný autozóm. U určitých druhů hmyzu a rostlin, které mají tzv. meiotický tah, ovšem může vzniknout životaschopné potomstvo.
Pericentrické inverze
Tyto inverze vedou ke změně v počtu ramen, ale nikoli v počtu chromozómů. Počet ramen chromozómu závisí na pozici centromery. Pokud se centromera nalézá na konci, pak má chromozóm jen jedno rameno. Pokud je uprostřed, pak má ramena dvě. Tento druh inverze může změnit akrocentrické chromozómy na metacentrické. Tato inverze odlišuje i křečky Neotoma od křečků Peromyscus.
Paracentrická inverze
U tohoto typu inverze není zahrnuta centromera. Nevyskytuje se příliš často, ale vědci se domnívají, že se objevuje u několika druhů netopýrů, zajíců a primátů.
Rozsáhlé změny
Za jistých podmínek, daných silným tlakem prostředí, může dojít k rozsáhlým změnám uspořádání genů, které mohou vést ke vzniku výraznějších variant, a přispět tak k přežití. Tyto změny mohou být rychlé, dostane-li se organismus do nové adaptační zóny (tzv. „regulační model“). Takovéto změny v uspořádání vědci našli u čeledi hlodavců přizpůsobených životu pod zemí – slepců (Spalax).
Při těchto rekombinacích je informace přeskupena, ale srovnání mezi proužkováním chromozómů u příbuzných druhů s různou chromozómovou strukturou ukazuje, že informace je stále stejná. Je jen jinak uspořádaná. Navíc platí, že typ změny v uspořádání, k němuž dochází u různých živočichů, je specifický pro určitou skupinu, a jeden typ přeskupení se tedy nemusí nutně vyskytovat u jiné.
AUTOR:

Tento článek je převzatý z knihy Genesis konflikt od prof. Waltera J. Veitha, doktora zoologie, mezinárodně uznávaného vědce, přednášejícího v mnoha zemích Afriky, Evropy, Ameriky a Austrálie. Profesor Veith věří, že evoluce neposkytuje uspokojivé vysvětlení našeho původu. Jeho kniha Genesis konflikt, stejně tak i série videí, která předkládá myšlenky z této knihy, jsou k dispozici v našem eshopu (nebo ZDARMA ke shlédnutí).
Prof. Dr. Veith v této knize prezentuje své studium původu života. Jeho přírodovědecká vášeň ho přivedla k radikálnímu životnímu přehodnocení jeho dřívějšího evolučního pohledu na svět. Upozorňuje na mnohé omyly i neznalost mnoha důležitých objevů. Mýty v dané oblasti systematicky vyvrací a nahrazuje alternativou kreacionistického výkladu přírodních jevů.
Další články ze série: Původ života a rozmanitosti
- Prvotní atmosféra
- Problém kyslíku
- Organické molekuly
- Aminokyseliny
- Nukleová kyselina - stvořil ji "nový bůh"?
- Přírodní výběr a tvořivá síla
- Přírodní výběr
- Teorie Ernsta Haeckela o vzniku vyšších forem života
- Je hypotéza Gastraea životaschopná?
- Mechanismy pro změnu
- Variace již obsažené v genofondu
- Reprodukční výměna
- Transpozibilní elementy
- Rekombinace chromozomů
- Biblická koncepce „druhů“ versus současné druhy
- Proč je tolik druhů?
- Proč je tolik druhů? – slovníček
- Distribuce po potopě
- Australský problém
- Evoluce: Zázrak zázraků
- Stvoření života ve zkumavce?


Komentujte