
Jak vznikla všechna ta rozmanitost, kterou vidíme v dnešním světě? Kolik „druhů“ bylo potřeba nalodit na Noemovu archu, aby se dala vysvětlit šíře a rozmanitost živočišné říše, která je dnes ozdobou naší planety?
Ve své knize Fundamentální biologie píše Frank Marsh:
“Ten, kdo si myslí, že (moderní) druhy zvířat a rostlin zůstávají stejné v po sobě následujících generacích, musí pouze prozkoumat přírodní záznamy, aby objevil svou chybu. Variace je jedním z nejvíce neměnných zákonů v biologickém světě.“ (1)
Podle Biblické knihy Genesis, ryby, ptáci, rostliny, suchozemská zvířata a lidé, pocházeli ze samostatných zákonů stvoření. Bible nazývá stvoření, která Bůh vytvořil jako “druhy” (Genesis 1:21). Později v Genesis čteme, že Noe přinesl do archy dva páry každého nečistého zvířete a sedm párů všech čistých zvířat. Přečtěte si více o čistých a nečistých zvířatech.
Jak se objevily druhy?
Podle L. J. Gibsona z výzkumného ústavu Geoscience Research Institute může mnoho druhů zvířat vzniknout několika způsoby. Kreacionisté věří, že Bible správně uvádí, že Bůh vytvořil “druhy” zvířat. Nicméně místo toho, aby Bůh vytvořil každý jednotlivý (moderní) druh, měly tyto druhy obrovskou vestavěnou možnost variace, která umožňovala velkou rozmanitost. V našem moderním klasifikačním systému máme tendenci klasifikovat zvířata, která jsou stejného druhu, ale mají různá označení nebo vlastnosti jako různé “druhy”.
L. James Gibson prováděl ve vztahu k těmto otázkám důkladný výzkum literatury týkající se těchto otázek a našel obrovský počet různých možností, jak lze existenci současných druhů vysvětlit. (2)
Hlavní mechanismy změny a původu druhů je možné shrnout následovně:
1) Variace vytvářené šlechtěním na základě vrozeného potenciálu pro změnu: příkladem jsou nejrůznější vyšlechtěná plemena psů, koček, domácího skotu, holubů a drůbeže. U některých přirozeně se vyskytujících druhů se projevují podobné rozdíly v postupné změně v populaci nebo druhu ve vztahu ke směru nebo orientaci určitých faktorů prostředí, například řeky, horského pásma, severo-jižního pásma lesů, nadmořské výšky apod. Gradient změny je pozorovatelný například u americké užovky červené (Elaphe guttata), u níž se jednotliví jedinci liší často barvou i počtem šupin. Dalšími příklady vestavěných variací jsou obecně variace v barvě, hustotě srsti, tvaru a velikosti těla a změny související s ročními obdobími.
2) Ztráta genetického materiálu: ztráta genetického materiálu (nebo deaktivace projevu genu) vedla k tzv. speciaci (vzniku druhů). U ptáků je běžná ztráta schopnosti létat, a to zvláště u druhů žijících na ostrovech, kde by pro ně létání mohlo představovat jisté znevýhodnění – hrozí jim totiž, že by je silný vítr mohl odnést na volné moře, odkud by se už nedokázali vrátit.
Příbuzné druhy si však často schopnost létat zachovaly. Jako příklad si můžeme vzít indonéského chřástala (obývá subtropické a tropické nížinné pralesy a bažiny), kormorána galapážského se zakrslými křídly, bernešky havajskéi, ztrátu očí u slepých jeskynních ryb nebo řady druhů hmyzu obývajících jeskyně. Ztráta očí u havajských švábů, obývajících jeskyně, ukazuje na mimořádně rychlou proměnu (v řádu několika měsíců), k níž dochází u těchto živočichů poté, co začnou obývat nově vytvořené jeskyně. Je vysoce nepravděpodobné, že geny pro oči po tak krátké době zmutují či z genofondu zmizí; dá se tedy spíše předpokládat, že geny zodpovědné za rozvoj očí jsou v podmínkách panujících v těchto jeskyních deaktivovány. Vzhledem k tomu, že změny prostředí, jako například cyklus světla a tmy, změny teploty během ročních období, mají na geny přímý vliv a mohou vést k aktivaci a deaktivaci genů, je tato možnost vzniku rychlých a dokonce velice výrazných změn vysoce pravděpodobná.
Máme-li mluvit o standardním způsobu klasifikace, pak ztráta genetického materiálu vede ke vzniku nových druhů nebo rodů, ale nikoli vyšších kategorií. S ohledem na stávající znalosti principů fungování genomu a toho, jakým způsobem jsou aktivovány a deaktivovány geny, je možné pochybovat, zda v případě vzniku druhů skutečně dochází ke ztrátě genetické informace. Je dosti pravděpodobné, že geny jsou pouze deaktivovány, neboť prostředí nevyžaduje vývoj daného rysu. Musí přirozeně existovat mechanismy, které dokáží deaktivovat dokonce i strukturní geny, kódující morfologické rysy, pokud toho bude zapotřebí. Tyto změny je možné považovat za příklady mikroevoluce, ale ve skutečnosti se v nich odráží jen standardní aktivita genomu. Neexistuje proto žádný oprávněný důvod řadit ryby obývající jeskyně, u nichž se nevyvinuly oči, do nového druhu jen na základě toho, že vypadají jinak. Středoamerická ryba tetra mexická (Astyanax fasciatus) má dvě formy: slepou jeskynní a říční, jež má oči. Obě formy se mezi sebou bez problémů kříží, musí se proto jednat o tentýž biologický druh. Přečtěte si více o přírodním výběru.
3) Hybridizace. Většina hybridů není životaschopná, protože ztrácejí plodnost – zvláště u savců. Některé skupiny živočichů (taxony) však mají sklon k hybridizaci a mohou dát vznik životaschopným druhům živočichů (například ryb nebo hmyzu) nebo celé řadě rostlin. Vznikají také hybridy skotu (sedm druhů rodu Bos – tur – může podléhat hybridizaci). Je možné vyšlechtit hybridy i mezi rodem tur a americkými bizony (Bison bison). Hybridy mohou vznikat také mezi koňmi a zebrami, velbloudy a lamami, levharty a jaguáry, lvy a tygry, delfíny a kosatkami černými (jiné rody) nebo různými rody hadů. Byly vyšlechtěny i hybridy mezi ovcemi a kozami, ačkoli v tomto případě byla embrya uměle zkombinována a poté implantována do dělohy náhradní matky.ii Genetická informace může být pro vznik reprodukční izolace rychle reorganizována několika způsoby, ale vzhledem k tomu, že nemusí nutně docházet k její ztrátě nebo vzniku nové informace, tyto izolované formy nepředstavují nový „druh“ (v biblickém chápání), nýbrž pouze příbuzný druh (species) téhož „druhu“ (rodu nebo čeledi), který je nyní reprodukčně izolován. V minulosti mohly proto různé druhy v rámci téhož rodu (případně i čeledi) náležet do téhož „druhu“.
4) Změny ve struktuře a počtu chromozomů: chromozomy jsou klasifikovány na základě umístění centromery (zúžené místo na chromozomu, kde jsou sesterské chromatidy po replikaci navzájem spojeny). Když je centromera uprostřed chromozómu a obě ramena mají stejnou délku, je chromozóm označován jako metacentrický chromozóm. Pokud se centromera nalézá poblíž konce chromozomu, pak se chromozóm nazývá akrocentrický. Veškeré změny ve struktuře chromozómu je možné odhalit pomocí zvláštního proužkovacího barvení.
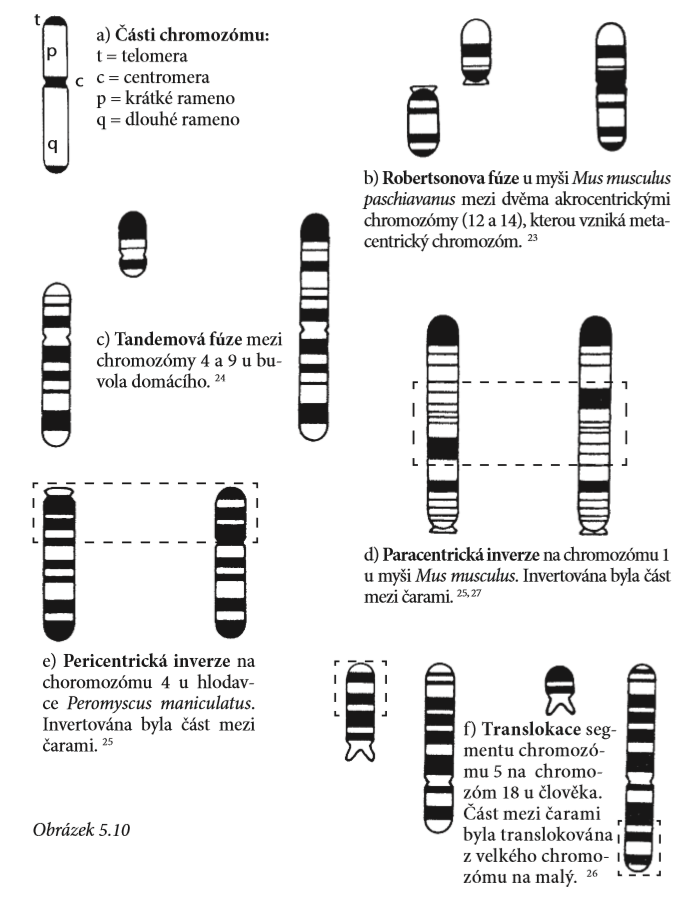
Změny na chromozomech mohou zahrnovat změny v počtu chromozomů, počtu ramen i jiné změny vyvolané translokací (přesunem segmentu chromozomu do jiného místa), vynechání, duplikace, inverze i drastické změny uspořádání. Chromozómy mohou občas fúzovat (spojovat se), přičemž se vytvářejí mnohem delší chromozómy, nebo se mohou v centromeře rozdělit, čímž se vytvoří dva kratší chromozómy. Jedna takováto změna uspořádání je označována jako Robertsonovo přeuspořádání a je výsledkem buď splynutí dvou centromer do jedné, nebo rozdělení jedné centromery na dvě. Tandemová fúze je naopak splynutí dvou chromozómů, při němž se jeden konec chromozómu spojí s koncem nebo centromerou jiného chromozómu (viz obrázek 5.10). Při těchto rekombinacích je informace přeskupena, ale srovnání mezi proužkováním chromozómů u příbuzných druhů s různou chromozómovou strukturou ukazuje, že informace je stále stejná. Je jen jinak uspořádaná. Navíc platí, že typ změny v uspořádání, k němuž dochází u různých živočichů, je specifický pro určitou skupinu, a jeden typ přeskupení se tedy nemusí nutně vyskytovat u jiné.
Tyto změny ve struktuře vytvářejí různé charakteristiky, které vědci často používají jako kritérium nového druhu. Ale bližší pohled na genetiku ukazuje jiný příběh. Zjistěte více o tom, jak funguje každý z následujících procesů: Fúze chromozomů, Robertsonova fúze, Tandemová fúze, Pericentrická inverze, Paracentrická inverze, Translokace a rozsáhlé změny.
Dá se proto obecně říci, že genom organismů je obdařen obrovským potenciálem pro tvorbu variací. V podmínkách zvýšeného tlaku prostředí nebo dostane-li se organismus do nových adaptačních zón, příp. při nízkém selekčním tlaku, se projeví zabudované mechanismy pro vytváření rychlých změn a obrovských variací. Organismy musely být tímto potenciálem pro změnu obdařeny již od samotného počátku a byly stvořeny ve stejnou dobu.
Skutečnost, že kmeny se ve fylogenetických dokladech objevují náhle, naznačuje, že tyto skupiny neměly společného předka. Nepřítomnost předků v níže položených vrstvách tedy není způsobena nedostatečným množstvím fosilních dokladů, ale naznačuje, že různé skupiny se vytvořily odděleně. Kdyby všichni tito živočichové patřili k biblickým „druhům“ a byli obdařeni jedinečným genofondem, pak bychom zřejmě mohli čekat, že se u nich budou projevovat určité prvky společného plánu. Genomy by však nevytvářely kontinuální řadu pokročilých genetických rysů od nejjednodušších po nejsložitější, protože všechny „druhy“ by byly jedinečné, ačkoli každá skupina by obsahovala potenciál ke tvorbě variací.
Společný předek tudíž neexistuje a neobjeví se ani ve fosilních záznamech. Přesně to fosilní záznamy ukazují (žádní společní předkové se nikdy nenašli). Vědci byli zklamáni, neboť ani fylogenetické studie nedokázaly odhalit společný původ všech organismů. Není divu, že někteří fylogenetikové uvažují o tom, že by se mělo upustit od snahy najít kořeny fylogenetického stromu, neboť molekulární fylogeneze organismů je plná genetických anomálií. Tvrdí, že laterální transfer (přenos) genů situaci neuvěřitelně zkomplikoval.(3) Je však možné, že se pouze dívají na nesprávné místo a přehlížejí takové řešení, které odpovídá nejen datům, ale i Písmuiii.
Fantastická řada mechanismů, které umožňují tvorbu změn a variací, se hodí zvláště v měnícím se prostředí nebo v situaci, kdy stabilizující selektivní tlaky jsou slabé – což byla přesně situace po potopě. Nově vytvořené adaptační zóny v době po potopě byly ideální pro rychlou adaptační radiaci (pomocí vrozeného potenciálu pro tvorbu genetických variací) a také soupeření o životní prostor (stanoviště) muselo být zpočátku slabé, což vedlo k nízkému stabilizujícímu výběrovému tlaku. Tyto podmínky mohly proto rychle vyvolat změny ve formě i struktuře, dokud se populace nestabilizovaly. Jedním z mnoha příkladů jsou velcí savci se svými extrémními formami v podobě mamutů a šavlozubých tygrů.
Vezmeme-li navíc v úvahu tento obrovský potenciál ke změně a zjevné vztahy dokonce i mezi druhy se zcela odlišným počtem chromozómů, je možné představit si situaci, kdy relativně malý počet „druhů“ dokáže vysvětlit i vznik velkého počtu druhů (species) ve velice krátké době. Ani pro lidi, kteří věří v pravdivost biblického příběhu o Noemově arše, již nemusí být představa nutnosti umístit všechny druhy živočichů do archy tak nemožná. Do archy nemusely nastoupit všechny druhy současných živočichů, jen zástupci vyšší taxonomické kategorie. Například by stačilo, aby antilopy byly zastoupeny jednou z variant, nikoli všemi, které jsou dnes rozděleny do různých druhů.
Velice zajímavou ilustrací jsou psovité šelmy. Vlci a psi rodu Canis mají 78 chromozómů, zatímco lišky mají mezi 38–78 chromozómy. Jednotnost v počtu chromozómů u psů a vlků může být způsobena širokou mírou křížení, zatímco lišky žijí v malých rodinných skupinách a na menších teritoriích, proto u nich přetrvává nové uspořádání. A tak třebaže počet chromozómů u mnoha psovitých šelem může být nižší než 78, které nalézáme u vlků, chromozómy druhů s menším počtem chromozómů jsou delší a proužkování ukazuje, že obsahují stejný základní genetický materiál. Delší chromozómy jsou tudíž výsledkem fúzí, které změnily počet chromozómů a informaci reorganizovaly. Přesto se jedná o stále stejnou genetickou informaci. Pokud bychom tedy za „druh“ považovali úroveň čeledi psovití (Canidae), pak již důsledky pro počet potřebných živočichů nezbytných pro vytvoření všech současných variací nejsou tak děsivé, jak se mnozí obávají. Velká proměnlivost forem a struktury, kterou vidíme u všech divokých psovitých šelem, není o nic větší než počet variací, jež dokázali lidé vyšlechtit u psa domácího. Všichni příslušníci čeledi psovitých tedy mohli původně náležet k témuž „druhu“.
Potenciál pro změnu v živočišné říši zcela jistě existuje, přesto existují i určité bariéry, které není možné překročit. Máme spoustu důkazů o tom, že všichni psovití museli původně náležet k jednomu „druhu“. V Kanadě se mezi západem a středem země objevuje variace ve velikosti kojotů. Větší kojot obývající střední Kanadu se občas kříží s vlkem a menší kojot ze západu se bez problémů kříží s jinými kojoty. Je proto pravděpodobné, že náleželi ke stejnému „druhu“.
Totéž platí dokonce i pro tzv. jedinečné psovité šelmy, jako je australský dingo, kterému hrozí vyhubení, a to nikoli proto, že by jeho území zabíral jiný živočich, ale protože jeho genofond je postupně pohlcován psem domácím, se kterým se snadno kříží.
S genomem mušky octomilky (Drosophilla) jsou vědci schopni manipulovat do takové míry, že to dle názoru některých odborníků přesahuje hranici, které dosáhly veškeré [předpokládané] evoluční události v dějinách Země. Třebaže byly vytvořeny i velice bizarní formy, nikdy nebyla překročena hranice, kterou představuje „octomilka“. Stejně tak muselo dojít k velikým změnám v důsledku přeuspořádání chromozómů, k němuž nejspíše od stvoření došlo, přičemž časový rámec odpovídá krátké chronologii. Je proto možné si představit, že ke změnám, jež vedly ke vzniku onoho velkého počtu dnešních druhů, mohlo dojít velice rychle. U současných živočichů byla identifikována řada chromozómových homologií, které jsou pravděpodobně pouze výsledkem fúze chromozómů, jako je tomu v případě klokanů, u nichž je možné rozdíly mezi jednotlivými druhy vysvětlit na základě Robertsonových fúzí. Rozdíly mezi jednotlivými druhy hmyzožravců, netopýrů, primátů, mořských savců, hlodavců, králíků, zajíců a kopytníků je také možné vysvětlit jako důsledek přestavby genů.(4)
Podobnost mezi genetickými vazbami ovšem nemusí vždy odrážet blízké vztahy; někdy se v nich odrážejí podobnosti v designu, založené na funkčních požadavcích. Například geny pro konkrétní systémy enzymů jsou často umístěny na chromozómech, které mají u různých druhů podobné proužkování.(5) Podobnosti je proto možné vysvětlit také na základě funkce, nikoli původu. Podobné vzorce genetických vazeb u koček a lidí jsou téměř totožné jako při srovnání lidí a šimpanzů (6). Takto je také možné vysvětlit podobnosti v chromozómech lidí a lidoopů. Zajímavé je, že karyotypiv člověka se zdá být nejblíže k primitivnímu stavu, což nepodporuje hypotézu o tom, že lidoopové byli předchůdci člověka.(7)
Závěr
Žádný model původu druhů nemůže poskytnout všechny odpovědi, zvláště pokud si uvědomíme, že naše znalosti mnoha biochemických a genetických mechanismů jsou dosud značně neúplné. Kreacionistický model ovšem nabízí mnoho realistických odpovědí na otázky, které nás ohledně původu a rozmanitosti života trápí.
Najdou se oblasti, kde nám nedostatek znalostí bude muset nahradit víra, ale totéž platí i o evolučním modelu. Obě dvě hypotézy v konečném důsledku vyžadují víru. Otázka, kterou si musí každý z nás položit, tedy zní: která hypotéza si žádá víry více?
Přečtěte si také o distribuci zvířat po potopě.
Poznámky:
i špatní letci, několik vyhynulých příbuzných druhů nelétalo vůbec – pozn. překladatele
ii V tomto případě se nejedná přímo o hybrid, ale o tzv. chiméru. U chiméry si každá sada buněk zachovává svou vlastní druhovou identitu (genetickou informaci), zatímco hybrid je „přechodná“ forma mezi rodičovskými druhy. Chiméra má tedy čtyři rodiče, zatímco hybrid dva. – pozn. překladatele
iii Bibli – pozn. recenzenta
iv Soubor všech chromozómů v jádře buňky. – pozn. recenzenta
Odkazy:
1. Frank Marsh, Fundamental Biology (1941):101.
2. GIBSON, L. J. Chromosomal Changes in Mammalian Speciation: A Literature Review. In Origins. 1984, sv. 11, s. 67–89.
3. DOOLITTLE, W. F. Phylogenetic Classifi cation and the Universal Tree. In Science. 1999, sv. 284, s. 2124–2128.
4. GIBSON, L. J. A Creationist View of Chromosomal Banding and Evolution. In Origins. 1986, sv. 13, s. 9–35.
5. LALLEY, P. A. / McKUSICK, V. A. Report of the Committee on Comparative Mapping. In Cytogenetics and Cell Genetics. 1985, sv. 40, s. 536–566.
6. O’BRIEN, S. J. / NASH, W. G. Genetic Mapping in Mammals: Chromosome Map of Domestic Cat. In Science. 1982, sv. 216, s. 257–265.
7. YUNIS, J. J. / PRAKASH, O. The Origin of Man: A Chromosomal Pictorial Legacy. In Science. 1982, sv. 215, s. 1525–1530.
AUTOR:

Tento článek je převzatý z knihy Genesis konflikt od prof. Waltera J. Veitha, doktora zoologie, mezinárodně uznávaného vědce, přednášejícího v mnoha zemích Afriky, Evropy, Ameriky a Austrálie. Profesor Veith věří, že evoluce neposkytuje uspokojivé vysvětlení našeho původu. Jeho kniha Genesis konflikt, stejně tak i série videí, která předkládá myšlenky z této knihy, jsou k dispozici v našem eshopu (nebo ZDARMA ke shlédnutí).
Prof. Dr. Veith v této knize prezentuje své studium původu života. Jeho přírodovědecká vášeň ho přivedla k radikálnímu životnímu přehodnocení jeho dřívějšího evolučního pohledu na svět. Upozorňuje na mnohé omyly i neznalost mnoha důležitých objevů. Mýty v dané oblasti systematicky vyvrací a nahrazuje alternativou kreacionistického výkladu přírodních jevů.
Další články ze série: Původ života a rozmanitosti
- Prvotní atmosféra
- Problém kyslíku
- Organické molekuly
- Aminokyseliny
- Nukleová kyselina - stvořil ji "nový bůh"?
- Přírodní výběr a tvořivá síla
- Přírodní výběr
- Teorie Ernsta Haeckela o vzniku vyšších forem života
- Je hypotéza Gastraea životaschopná?
- Mechanismy pro změnu
- Variace již obsažené v genofondu
- Reprodukční výměna
- Transpozibilní elementy
- Rekombinace chromozomů
- Biblická koncepce „druhů“ versus současné druhy
- Proč je tolik druhů?
- Proč je tolik druhů? – slovníček
- Distribuce po potopě
- Australský problém
- Evoluce: Zázrak zázraků
- Stvoření života ve zkumavce?


Komentujte